Fisher's run-away sexual selection process
is a mechanism Fisher put forward to answer one of the big sex questions (go to the end) - the apparently excessive ornamentation seen (usually among males) in nature. See also....- 1012.pdf: Is the peacock merely beautiful or also honest?
- 49806.pdf: Evolution of multiple sexual pref. by Fisher's runaway process
- Runaway ornament diversity caused by Fisherian sexual selection - Pomiankowski and Iwasa paper
- 1991Nature350-332.pdf: Kirkpatrick and Ryan, Nature, 1991 -- READ THIS MARCUS
- NB. at http://bio.research.ucsc.edu/~barrylab/classes/animal_behavior/BOX_3_1.HTM:
there's a picture like the one Maciej Wojnar drew at FestivalOfDoubt, which nicely explains the Fisherian run-away process of sexual selection.
HERE'S AN IDEA :
maybe it can be even simpler: the Fisherian argument seems to work for any assortment mechanism. For example: if some members of a species prefer to aggregate at some (essentially random) location, and that's where they find mates, their offspring will tend to do the same. Compare this to non-aggregating members, most of whom may indeed produce non-aggregating offspring, but some of who will mate with aggregators and have aggregator progeny. So aggregators can come from ... you get the picture. Arbitrary aggregators flourish at expense of rational generalists.... Uber-biologist Jeff "rivet gun" Shima on sea stuff: - apparently there are these brittlestars (odd starfish things), that only use on the blue sponges as 'habitat' to hang-out in, even though it would appear there are lots of other habitable sponges around. The authors searched for obvious attributes of these blue sponges that they felt might impact on brittlestar fitness, and concluded there was no good reason for it... but they speculate that the preference for blue sponges was a good way for males and females to find one another (and mate)--because once they choose a sponge, they seem to have very limited mobility. So in Fisher's world, maybe it was just that females who hung around blue sponges would meet males who hung around the same, and that variants who didn't might do worse... So we wonder whether there's a Fisherian story here, because it sounds eerily familiar. The red-breastedness of male birds is their phenotype, and an "extended" phenotype of blue-loving brittle-stars is blue-brained-starfish and blue sponges in the same place. The argument's even simpler than the usual Fisher one. We don't need separate genes for "liking X" and "expressing X": just need "go to X" (where X is essentially arbitrary) for both sexes. But it seems too simple now... it's just assortment. MarcusEvolutionary game theory
Let's say we have an "aggregator" gene (A), that causes the bearer to aggregate somewhere to some extent - for example it could be a "go to the blue sponges at mating time" gene. The "somewhere" is going to be completely arbitrary, for argument sake: there is absolutely no fitness gain or loss for individuals with A genes over the other guys (let's call them generalists, " G ", but we don't need a gene for that - it's just the lack of an A gene). Say a proportion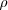 of the population currently carries the A gene, for both sexes.
Crude model of sexual haploid reproduction: one of the two parents' alleles is chosen at random to go into the offspring. Eg. if a G mates with an A, we get a GA combination so G half the time and A half the time. But if a G mates with another G, the offspring is definitely a G, and similarly two A 's can only make an A.
Assume that all females have the same number of offspring per "generation".
of the population currently carries the A gene, for both sexes.
Crude model of sexual haploid reproduction: one of the two parents' alleles is chosen at random to go into the offspring. Eg. if a G mates with an A, we get a GA combination so G half the time and A half the time. But if a G mates with another G, the offspring is definitely a G, and similarly two A 's can only make an A.
Assume that all females have the same number of offspring per "generation".
Offspring of generalist females
A female G mates with a male G with probability (makes GG), and mates with a male A with probability (giving GA). Thus offspring is G with probability
(makes GG), and mates with a male A with probability (giving GA). Thus offspring is G with probability 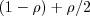 and A with probability
and A with probability 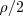 .
So the G females (current density of all females) provide a proportion
.
So the G females (current density of all females) provide a proportion 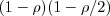 G individuals to the next generation, and
G individuals to the next generation, and 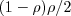 A 's.
A 's.
Offspring of fussy (aggregator) females
If there was no aggregation, a female A would mate with an A male with probability, but we assume this is higher due to aggregation. Introduce an aggregation coefficient (exactly like a "relatedness" on in kin selection models), 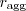 . This is a control knob for cranking the aggregation (assortment) up or down.
If
. This is a control knob for cranking the aggregation (assortment) up or down.
If 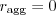 everything's the same as for generalists and a female A mates with a male A with prob . If
everything's the same as for generalists and a female A mates with a male A with prob . If 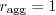 a female A will only ever come across (mate with) a male A (prob
a female A will only ever come across (mate with) a male A (prob 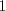 ). In between we transition linearly: a female A mates with a male A with prob
). In between we transition linearly: a female A mates with a male A with prob 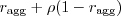 , and with a male G with probability 1 minus that.
So plugging that in, a female A will have offspring that are A with probability
, and with a male G with probability 1 minus that.
So plugging that in, a female A will have offspring that are A with probability 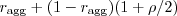 , and G with probability 1 minus that. And female A 's happen with probability in the current generation.
, and G with probability 1 minus that. And female A 's happen with probability in the current generation.
The next generation
Putting those together we can add up the frequency of A in the next generation, which (after simplifying) gives![\[ \rho^\prime \; = \; \rho + \frac{r_\mathrm{agg}}{2} \rho (1-\rho) \]](/foswiki/pub/Groups/AI/FisherianRunaway/latexceb4b7ccb77a2c05adb7fc9d44dc2da7.png)
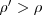 for any
for any 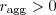 . So any assortment beats no assortment, and more assortment is always a Good Thing (meaning it will evolve if it can).
Presto! Marcus Frean
. So any assortment beats no assortment, and more assortment is always a Good Thing (meaning it will evolve if it can).
Presto! Marcus Frean
Maciej Wojnar says:
"That's a very interesting case, and I think that this is analogous to a Fisherian runaway. In normal sexual reproduction, a gene that is "fair" has 0.5 + 0.5p chance of being passed on to the next generation (where p is the proportion of the population that has the gene). This kind of reproduction does not alter the frequency of the gene in the population. I think that a Fisherian runaway occurs when a gene (or genes) cheats the system. If a gene can figure out how to make the individual more likely to mate with another individual that carries the same gene, then it will get a higher proportion of offspring with that gene and it will increase in frequency in the population. Because of this, the gene can significantly decrease the fitness of the individual and still spread in the population. I think that you hit the nail on the head when you compared it to assortment. Only it's better to think of it as gene assortment rather than individual assortment since it's the genes that are benefiting at the cost of the individual (the other genes). In the standard Fisherian runaway, the genes for big peacock tails are likely to to be found in peahens that like big tails (because their mothers probably liked big tails and so probably mated with a peacock with a big tail); similarly the genes for liking big tails are likely to be found in peacocks with big tails (because their fathers probably had big tails and so probably mated with a peahen that liked big tails). So in this case, two different genes cooperate together to assort themselves at the expense of the individual. In the case of the starfish, there is only a single "cheating" gene that uses an external cue to assort itself at the expense of the individual. This seems pretty plausible to me. I don't think that you can extend the Fisherian runaway to different species because, as I understand it, the engine driving it is the gene trying to skew the 0.5 + 0.5p chance of being passed on to the next generation."
Joseph Bulbulia Your thought: runaway selection doesn't require signals. Objection: Sex isn't elaborating it. It's natural selection operating through density dependent effects in both sexes, i.e. there is no variation among the sexes for selection to work with. If there is no signal relevant to choice, then sexual discrimination isn't causing anything. Rather, the dynamic you describe emerges from selection acting on both sexes, equally.
Marcus Frean : True, there's only the one gene and it has the same effect on both sexes. But it's still sexual selection in the sense that there's an essential sexual dichotomy (ooo!) there: females all get the same number of offspring, but males don't, just as in the picture, and this is what makes it work. So sex is still crucial. Is it still sexual "selection"? Well, what's the difference between mating with the peacock because he's next to the pretty tail, and mating with the peacock because he's next to the blue sponge? ("Hey, nice sponge you've got there, big boy!")Worry: ...in the signal instances, we can see how sex causes a trait to runaway. In the spatial assortment case we can't. What would it mean to not move from where you are placed, and to evolve to increasingly do this?
Marcus Frean : I wasn't thinking of it quite this way. Both sexes are at least partially mobile. I don't imagine anyone evolving not to move. Rather, you can evolve either to move in a generalist "unfussy" way, or you can move in a "fussy" way, preferring some places over others for not obvious fitness gain. And this latter tendency seems capable of running away. If it's "true" (whatever that means), why doesn't it happen more often then? Sure, it's daft, but it's no MORE daft (and a good deal less complicated to 'implement') than peacock's tails etc...
More thoughts
Joseph Bulbulia Neat! By "grain of selection," I meant to notice that natural selection operates on variation in a population. The variation you imagine is arbitrary location preference. The question is thus: what evolutionary process is maintaining and elaborating evolutionary change in a population exhibiting variation in location preference? We imagine that sexual selection is just natural selection operating through mate choice. So the question is whether mate choice tracks the variation. We can imagine two cases: 1) mates track signals and 2) mates do not track signals. To take the former, we imagine that geographic preference itself is exogenous to mating decisions. The spatial preference trait makes mating more likely, independent of choice. It does so merely by throwing horny beasts together (at the limit we can imagine no partner discrimination -- it's 3am Saturday night, Courtney place. The gene sends you there).* The second case: beasts evolve partner preference for spatial preference traits. They evolve p-preferences for those that hold s-preference traits. We assume variation in both. This is clearly sexual selection, because p and s perferences co-evolve. There's evolution here to recognize the spatial trait, which favours and elaborates both recognition and location traits. And it is Fischerian because it looks like a runaway effect can kick in, assuming that being in a location is subject to degrees (the "here" shrinks?). But to show this case is interesting there must be some interesting difference in the relevant preferences, sexual and spatial, to the normal case of doing something stupid but eye catching and getting the girl... I'd rather see the exogenous sexual selection case defended. Your key insight is that we can get the sexual runway process that Fischer imagined without requiring discrimination or choice. Next step, I think, would be to show that this isn't an outlier, but rather something that is quite general. My hunch is that it has wide application to coordination games. *The exogenously selected story recollects a Seinfeld episode (which I'm misremembering) in which Jerry recommends to George that he live with this woman, because that's how breeders get animals to mate, just by putting them in the same cage. (Perhaps Jerry was warning George about this fact, as his girlfriend moved in with a younger athletic man with good hair.) What process causes the opportunism here? Sexual choice or just having sex? Some might have a hard time viewing this as sexual selection.JeffShima - This is all well outside of my expertise, but I do find it fascinating. I'm not sure if these are ideas that people learn in Evolution 101 because (sadly) I never took such a course. It would not surprise me at all though, if the ideas unfolding on your wiki are new (or least different enough from the way that folks have typically conceived the problems). I think we should discuss further over alcoholic beverages at some stage--I find that this helps my creativity greatly. Ultimately it would probably be nice to have some data/empirical evidence to draw upon (I'm thinking of those reviews I received for my Am Nat cum Eco Letters cum Ecology manuscript). I must confess a measure of ignorance about that brittlestar-sponge example--I haven't read the manuscript all that carefully--I'm not even certain the sponges are blue!--(and based on the reviews coming in, I'm likely to reject it for Oecologia)--but at least it gives some names and context to mythical assorting creatures. Joseph's point about this sort of mechanism possibly leading to evolution of creatures that don't move is an interesting one. Continuing with marine creatures as a hypothetical subject, most species produce offspring that (have the potential to) disperse widely, and then metamorphose into adults that hardly move. This is likely the ancestral trait, and to some degree its probably constrained by ancestry and trade-offs with of attributes of life-history. Interestingly, I did note that the brittlestar in this manuscript that I barely read was a "direct developer" (which means it lost the ancestral dispersive stage) for the most part (though the authors claim the the juveniles still disperse much more than the adults). This is perhaps a bit too tangential, but I wonder if, for example, one were to conduct a meta-analysis (not sure if data exist, but lets assume so for now), they might find that species that tended to exhibit strong patterns of "assortment" also tended to be ones with reduced (or lost) dispersal potential... I'm not sure there is a logical connection here--I'm mostly just wondering if there might be a way to bring in some empirical evidence/support/motivation... I do think I recall some speculation that marine creatures with greater dispersal potential (i.e., presumably with less loss of the ancestral trait) also tend to be generalists with respect to settlement habitats (e.g.,the color of sponges they like). The argument--if I'm remembering this correctly--is that cretures with high dispersal potentials "need" to be generalists, because the environment that they may end up in is just that much more uncertain... But perhaps the mechanism that you and Joseph describe is a plausible alternative... I also find this all quite interesting because (I think) it sets up a scenario whereby sympatric speciation might occur--I not sure if the mechanism you're arriving at is one that is already acknowledged in this context, but I do believe (based upon my very limited expertise in evolutionary questions) that there is still a bit of uncertainty and debate about how sympatric speciation could ever happen--and I think the assortment scenario is potentially one possible answer. Short answer: I think the idea is really cool.
Musical chairs simulation
Marcus Frean - I ran a simulation for the fun of it. Each creature has a genome consisting of a favourite spot in the range 0 to 1, and a tolerance around that favourite that it's prepared to put up with. "Spot" here could be an absolute point in space, or could mean "next to the blue sponges, wherever they are". It could be that the blue sponges are handy for other reasons (like eating), but that's beside the point here. I think the point is that "favourite" could refer to just about any bizarre and arbitrary conjunction of conditions. The simulation starts with everyone having tolerances of 100%: i.e. they don't care where they are at all. In each generation, the 100 creatures take up positions randomly but within tolerance of their favourite spot. All the females then get pregnant by the nearest male (this is starting to sound like musical chairs or something!!). Females then give birth to 2 offspring each. To make an offspring, toss a coin to decide which parent's genome to use. Mutate that genome slightly. Toss another coin to determine the sex. Then we kill all the parents and do it again, and so on for (say) 10000 generations. Here's what happens - red is early on, going blue as time goes by (the numbers are # generations). The y-axis shows general (top) to specific (bottom), the x-axis shows the arbitrary spot that's being favoured. Each point is the genome of an individual creature, of which there were 100. They clump up (despite periods of seeming to unclump). Note
They clump up (despite periods of seeming to unclump). Note - There's absolutely no fitness advantage or disadvantage associated with any particular place.
- There's no advantage in clumping up per se.
- Females mate with the nearest male: they don't have to be "close". So all females have the same 'basic' (meaning not-including-inclusive) fitness.
Joseph Bulbulia Suppose you could do this with time, explaining why my ex-misses never mated with me, even though we lived together. Question: "females all get the same number of offspring, but males don't" -- the averages must be the same. Still uncertain of the role sex plays here. Females mate with the nearest man, if there is no female in between. Men mate with the nearest female, if there is no a man in between. But I'm slow. Much hunch is that you don't need sex. The deeper point is has to do with assortment, and runaway elaboration on a trait, like you said in the first sentence. I like thinking about this a coordination problem. (While it's true the "women" will mate no matter what, there's a basin favoring proximity -- "all females mate w/ the nearest man", which again seems to me symmetrical.) But I'm probably missing something obvious.
Maciej Wojnar I think that Joseph is right that you will still see this aggregation taking place if the sexes are completely symmetrical. It would be interesting to modify the simulation so that each individual mates with its nearest neighbour and produces 1 offspring, thus eliminating any difference between the sexes (essentially eliminating the sexes while still having sexual reproduction). Also, as I was looking at the graph, it seemed to me that there would be massive overcrowding and competition for resources if all the "creatures" tended to congregate in one area. Perhaps adding a fitness cost proportional to the number of creatures in the same area to the simulation would be illuminating. Another thought occurred to me. Perhaps this kind of behaviour is widespread in nature, but the creatures only congregate during the mating season to mitigate the cost of clumping together and competing for the same resources.
Where/when/why
Females that like doing it in the elevator may be no more fit than those who do it anywhere, but the males that do it in the elevator are slightly fitter and that's apparently enough to reward elevator fussiness. Put another way, the gene "only ever do it in the elevator" should be propagated, because individuals with it have more offspring on average than those that don't. Marcus Frean - maybe it'd help to make a list of when to mate.| when to mate? | why that makes sense |
|---|---|
| when near other resources | meets two needs at once |
| when near other individuals | well, mating's easier! also (eg) females can select among the available males, based on signals |
| when some arbitrary condition is true | *It doesn't have to "make sense" in terms of the individual. For the gene, it solves a coordination problem* |
How old-hat / trivial is this?
Googling with "Fisher's runaway process aggregation" gives things like.................................... This must be related to assortative mating, although wikipedia (!) says"For some populations there are two different resources for which different phenotypes are optimum. Intermediates between these two phenotypes are less favorable. It is then favourable if the organisms can recognize mates that are optimized for the same resources as they are themselves. If mutations that make such recognition possible appear, these will be selected for."But that seems to go too far: don't need to recognise mates, and intermediates don't need to be any less favourable. This (below) looks close, but I don't think you even need "resources" to "over-aggregate" on (although that's a natural place to aggregate TO, I guess).: http://www.springerlink.com/content/n722653437464150/
"*Summary:* Resource-defence polygyny is a mating system that offers an opportunity for strong sexual selection. I express the degree of aggregation or clustering of females at resources at the time of mating using Lloyd's measures of microspatial aggregation, mean crowding ( m *) and patchiness ( P). Here, I show... (etc. etc.). As a result, females are selected to over-aggregate with respect to resources. The run-away process halts when single males cannot defend and maintain exclusive reproductive access to clusters of females as evidenced by extraharem copulations and paternity."


| I | Attachment | Action | Size | Date | Who | Comment |
|---|---|---|---|---|---|---|
| |
1012.pdf | manage | 417 K | 27 Nov 2008 - 17:08 | Main.marcus | Is the peacock merely beautiful or also honest? |
| |
1991Nature350-332.pdf | manage | 1 MB | 20 Jan 2009 - 13:45 | Main.marcus | Kirkpatrick and Ryan, Nature, 1991 |
| |
49806.pdf | manage | 1 MB | 27 Nov 2008 - 17:16 | Main.marcus | Evolution of multiple sexual pref. by Fisher's runaway process |